
Несмотря на значительное количество работ по технологии вермикомпостирования, сведений по экологии калифорнийского гибрида дождевого червя сравнительно мало. Нами проведены исследования некоторых экологических особенностей вермикультуры Eisenia foetida: получение биомассы взрослых особей, воспроизводство коконов и яиц при получении вермикомпоста из птичьего помета, фекалий, опилок и пищевых отходов, учет паразитов - личинок тахины или ежемухи Tachina radis и пораженных ими червей.
При изучении экологических особенностей дождевого червя выявлено, что на его пищевую и репродуктивную активность, получение биомассы влияют абиотические и биотические факторы, такие как температура, влажность, качественный состав структурообразователя, субстрата и наличие конкурентов и паразитов.
Влияние трофического фактора на культивирование дождевого червя Eisenia foetida и выход вермикомпоста. По данным наших исследований (табл. 1), за три месяца переработки субстрата в начале опыта средняя масса одной особи взрослых червей составляла 0,35 г, наибольший прирост средней массы червя на конец опыта обнаружен в варианте с использованием пищевых отходов (0,16 г) и птичьего помета (0,10 г). Средний прирост биомассы дождевого червя в варианте с использованием фекалий составляет 0,06 г. Опилки не содержат достаточного количества питательных веществ, поэтому черви на данном субстрате длительно существовать не могут. За срок 90 суток биомасса одной особи снизилась на 0,06 г.
Таблица 1 Влияние субстрата на получение биомассы дождевого червя
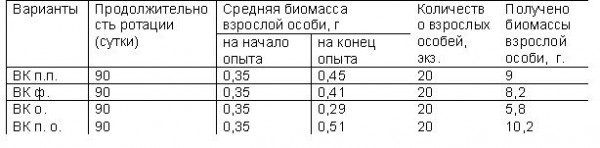
Для получения большей биомассы предпочтительнее использовать субстрат из пищевых отходов и птичьего помета.
Высокая активность размножения Eisenia foetida наблюдалась в вариантах с использованием субстрата из птичьего помета и фекалий, где оплодотворение происходило через 7 суток. В варианте с использованием пищевых отходов и опилок оплодотворение происходило реже, особенно при пониженной температуре (от +10 до +190С), через 11-15 суток. В исследованиях культивировали молодых и взрослых особей дождевого червя, у которых репродуктивная активность несколько различна (под молодыми особями мы имели в виду тех червей, которые стали половозрелыми от ювенильного возраста через 3 месяца, а под взрослыми – тех, возраст которых больше года.). Выход неполовозрелых особей у популяции взрослых червей выше, чем у молодых. При благоприятной температуре (от +20 до +250С) молодые особи способны выплодить от 1 до 3 экземпляров из одного кокона, а в холодный период - 1, реже - 2 (табл. 2,3). В сравнении с литературными данными это на 70% меньше, чем в самых благоприятных условиях, где выход неполовозрелых особей из одного кокона составляет от 9 до 20 экземпляров. По лабораторным исследованиям А.Ж. Барне (2002), количество зародышей в коконе составляло 2-3 экземпляра, но из каждого кокона чаще вылуплялась одна особь, что подтверждается нашими исследованиями.
Таблица 2 Влияние субстрата на выход коконов у дождевого червя, экз.

Таблица 3 Влияние субстрата на выход неполовозрелых особей из кокона у дождевого червя, экз.
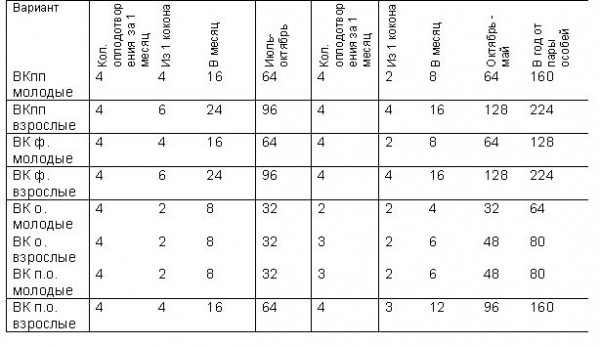
В условиях нашего опыта расчет структуры популяций молодых и взрослых особей в субстрате из опилок и фекалий показал, что выход молоди ниже, чем в остальных исследуемых субстратах. Вероятно, это связано с малой скоростью эмбрионального и постэмбрионального развития. В субстрате из птичьего помета и фекалий у молодых и взрослых особей выход коконов и молоди был выше. Это свидетельствует об оптимальности условий в данном субстрате.
В варианте с использованием птичьего помета (табл. 4) выход вермикомпоста к субстрату составил 98%. На втором месте по выходу вермикомпоста к субстрату стоит вермикомпост, полученный из фекалий, - 96%. Фекалии содержат меньшее количество питательных элементов в сравнении с птичьим пометом, червю же для полноценного питания необходимы сахара, белки, углеводы и другие питательные элементы. В варианте с использованием пищевых отходов развился плесневой гриб, что отрицательно повлияло на пищевую активность червей, с этим связан низкий выход вермикомпоста к субстрату -91 %. Выход вермикомпоста в варианте с использованием опилок составил всего 78%, что значительно ниже остальных вариантов.
Таблица 4.Влияние субстрата на получение вермикомпоста
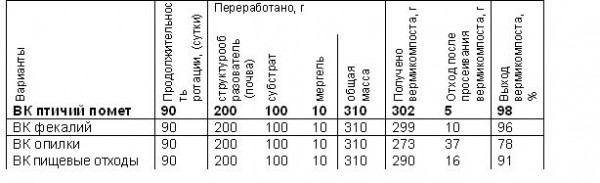
Для получения большей биомассы дождевого червя можно использовать субстрат из пищевых отходов, но этот вид отходов подвержен развитию плесневых грибов, что влияет на выход вермикомпоста. Поэтому пищевые отходы, фекалии и опилки необходимо смешивать с другими отходами.
Таким образом, оптимальные условия в субстрате из птичьего помета значительно влияют на пищевую и репродуктивную активность, а также получение большей биомассы Eisenia foetida и вермикомпоста.
Влияние температурного режима на выход неполовозрелых особей Eisenia foetida
Температурный режим повлиял на репродуктивную активность Eisenia foetida (рис. 1). Культивирование при температуре +100С показало наибольший выход неполовозрелых особей у взрослых червей в вариантах с использованием пищевых отходов и птичьего помета. В вариантах с использованием опилок и фекалий выхода молоди не наблюдалось. При температуре от +10 до +160С выход неполовозрелой особи был не во всех коконах, значительно ниже их выход наблюдался у молодых особей в варианте с использованием опилок и фекалий. При повышении температуры до +180С обнаружен выход молоди во всех коконах, но в варианте с использованием опилок наблюдался выход только по одной неполовозрелой особи из кокона, что снижает скорость вермикомпостирования. В остальных вариантах выход неполовозрелых особей из одного кокона составлял 2 экземпляра. При оптимальной температуре (от +20 до +250С) в варианте с использованием птичьего помета и пищевых отходов выход молоди из одного кокона составлял 3 экземпляра.
Экологический оптимум дождевых червей составляет от +20 до +250С. При пониженных температурах (от+10 до +150С), вероятно, не происходит оплодотворения яйца или развития зародыша у большей доли коконов, что значительно замедляет процесс вермикомпостирования.
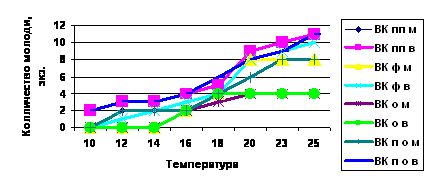
Рис. 1. Влияние температуры на выход неполовозрелых особей из коконов у Eisenia foetida.
Для активного размножения червей необходимо создавать температурный режим от +20 до +250С. При необходимости можно сохранить вермипопуляцию, снизив температуру до +100С.