ля нормального развития и роста рыбы, как и другие животные, нуждаются в определенном наборе питательных веществ. При составлении пищевых рационов необходимо учитывать пищевые потребности конкретного объекта выращивания и его физиологические особенности к усвоению компонентов, входящих в состав искусственных диет.
Кроме того, рыбы не могут синтезировать некоторые вещества и обязательно должны получать их с пищей. Короткий пищеварительный тракт рыб и низкая температура обитания препятствуют развитию обильной микрофлоры, которая у теплокровных животных в значительной мере обеспечивает организм витаминами. Необходимый для нормальной жизнедеятельности витаминный комплекс, в том числе и витамин С, имеющий большое значение для роста и развития, рыбы получают с пищей.
Потребности рыб в питательных веществах регулируются генетически обусловленным уровнем обмена веществ.По характеру потребляемой пищи (размерному составу и систематической принадлежности жертв) рыбы сильно отличаются друг от друга. По разнообразию потребляемой пищи рыбы стоят на первом месте среди других групп животных.
Питание в начальный период жизни - развитие в икринке и сразу после вылупления эмбриона – происходит за счет запасов желтка и жира в желточном мешке (эндогенное питание).
После перехода на внешнее питание, а также по характеру и разнообразию потребляемой пищи рыбы сильно отличаются. Приспособляемость к питанию определенными кормами не остается постоянной в течение жизни рыбы и меняется по мере роста и в связи с изменениями внешних условий обитания. Личинки практически всех видов рыб питаются зоопланктоном, и только по мере роста они переходят (за исключением зоопланктофагов) на другие пищевые объекты, т.е. спектр питания с возрастом изменяется. По характеру питания взрослых рыб делят на хищных и мирных.
Хищные рыбы (лососи, треска, щука, судак, сом) питаются в основном рыбой и в меньшей степени другой пищей.
Среди мирных рыб выделяют бентофагов, планктофагов и растительноядных.
Приведенное деление в некоторой степени условно, так как рыбы, отнесенные к той или иной группе, нередко потребляют и другую пищу. Сазан (карп) поедает как растительную, так и животную пишу, что позволяет отнести его к всеядным рыбам. Белый толстолобик помимо фитопланктона потребляет детрит (отмершие организмы гидробионтов), бактериопланктон, которого много в естественных водоемах и рыбоводных прудах. Каждый вид рыбы приспособлен к питанию определенным кормом, его органы чувств приспособлены к отысканию этого корма, ротовое отверстие - к захватыванию, кишечник - к перевариванию.
Потребность рыб в энергии
Для нормальной жизнедеятельности рыбы и обеспечения правильного обмена веществ необходимо обеспечить в первую очередь поступление в организм достаточного количества энергии. Обмен веществ представляет собой результат всех химических и энергетических превращений, происходящих в организме.
В отличии от птиц и млекопитающих энергетические потребности рыб невелики. Для прироста 1 кг массы рыбы в ее пище должно содержатся 4000-5000 ккал, а для сельскохозяйственных животных требуется 7000-9000 ккал и больше. это связано с тем, что у теплокровных много энергии уходит на поддержание температуры тела поэтому энергопротеиновое отношение комбикормов для рыб составляет7-10 ккал на 1 г белка, а животноводстве 15-20 ккал энергии.
В составе полноценных комбикормов для рыб учитывают количество энергии (перевариваемая энергия).
Потребность в протеине
Для нормального роста и развития гидробионтов необходимо определенного количество и соотношение основных питательных веществ. Для составления пищевых рационов необходимо учитывать пищевые потребности конкретного объекта выращивания и его физиологические особенности к усвоению компонентов, входящих в состав искусственных рационов. Потребность рыб в питательных веществах регулируется генетически обусловленным уровнем обмена веществ.
Основными питательными веществами корма являются протеин, жир, углеводы, минеральные вещества, витамины и другие биологически активные вещества, которые должны находиться в составе корма в соответствии с потребностью гидробионтов.
Протеин – основная часть живой материи, материал, идущий на построение тканей и органов в течение всей жизни. Понятие «протеин» включает в себя белковую (высокомолекулярные белковые вещества, полипептиды, олигопептиды, аминокислоты) и небелковую формы азота, различающиеся по качеству.
Белковые вещества обеспечивают рост и обновление тканей, участвуют в функционировании метаболизма, принимая участие в дыхании, пищеварении, в транспорте кислорода и переносе питательных веществ с током крови, в защите организма и размножении.
У наземных позвоночных выведение конечных продуктов азотистого обмена (ионов аммония) происходит посредством обезвреживания аммиака путем синтеза мочевины, мочевой кислоты и выведения их с мочой через почки. Пресноводные и морские рыбы относятся к аммониотелическим животным. Отличительной особенностью гидробионтов от наземных позвоночных является механизм выведения конечных продуктов азотистого обмена в виде легкорастворимых в воде азотистых продуктов – аммиака и мочевины, которые удаляются преимущественно внепочечным путем. Почти весь аммоний у рыб (более 90 %) выводится через жабры. Такая биохимическая особенность выведения конечных продуктов азотистого обмена гидробионтов сделала возможным функционирование метаболизма с преобладанием белкового обмена.
Следовательно, отличительными особенностями метаболизма водных гидробионтов от наземных животных являются:
1. Благодаря специфическим условиям водной среды обитания функционирует удобная и мало энергозатратная форма выведения конечных продуктов азотистого обмена в виде легкорастворимых в воде азотистых продуктов – аммиака и мочевины через жабры.
2. Высокая скорость роста. Рыбы переходят на внешнее питание при очень малых размерах и растут чрезвычайно интенсивно, достигая в сравнительно короткий период дефинитивной массы.
3. Короткий пищеварительный тракт, приспособленный для утилизации легкоусвояемой белковой пищи. Относительная длина кишечника большинства рыб-хищников меньше или равна длине тела. У зоопланктофагов, бентофагов и всеядных рыб превышение длины кишечника не более чем в 2 раза длины тела. Лишь у растительноядных рыб длина кишечника превышает длину тела в 4-10 раз. У наземных животных кишечник, как правило, по длине превышает тело в 10-30 раз.
4. Высокая доля участия белка в энергетическом обмене (свыше 60 %).
В результате особенностей белкового метаболизма для водных гидробионтов характерна высокая потребность в белке, которая превышает потребность у наземных животных. Например, для молоди лососевых рыб оптимальный уровень белка в корме составляет 45-55 %, для взрослой рыбы – 35-45 %. Для карповых рыб потребность в белке более низкая составляет 35-40 %. Стартовый корм для карповых рыб должен содержать высокий уровень белка (50-55 %).
Потребность в углеводах
Обсуждаются структура, роль метаболизм липидов в организме гидробионтов. Приводится оптимальное соотношение белка и жира в комбикормах для основных видов аквакультуры.
Углеводы являются менее значимыми питательными веществами в метаболизме рыб. Это объясняется тем, что большая часть гидробионтов не приспособлена ассимилировать углеводы кормовых объектов. Как правило, потребности в энергии удовлетворяется за счет липидного и белкового обмена. По питательности углеводы подразделяют на легкогидролизуемые и трудногидролизуемые углеводы. Первая группа представлена моносахаридами, дисахаридами (глюкоза, сахароза, фруктоза, рибоза, мальтоза и другие). Вторая группа состоит из сложных углеводов полисахаридов (крахмал, гликоген, хитин, целлюлоза). Из легкогидролизуемых углеводов наибольшее значение в питании рыб имеют пентозы и гексозы. Из трудногидролизуемых – олиго- и полисахариды.
Углеводный обмен у рыб складывается из 5 этапов:
1. Гидролиз поступающих с кормом полисахаридов до моносахаридов и всасывание их в кровь.
2. Образование и отложение в печени гликогена.
3. Расщепление гликогена в печени до глюкозы, образование в печени глюкозы из метаболитов жирового (глицерин) и белкового (аминокислот) обмена и поступления его в кровь.
4. Расщепление в клетках глюкозы до молочной и пировиноградной кислоты (анаэробный процесс) и дальнейшее окисление в цикле Кребса до углекислоты и воды (аэробный).
5. Выведение продуктов распада.
Углеводный обмен у рыб идет менее эффективно, чем у теплокровных животных. За счет низкого продуцирования инсулина углеводный обмен у многих видов рыб, особенно хищников, носит характер диабетического и если рыба получает избыток углеводов, развивается синдром перегрузки печени гликогеном, водянка брюшной полости, увеличение смертности. Максимальный уровень углеводов в корме для молоди лососевых составляет 30-35%. Для других рыб, особенно карповых и канального сома возможен более высокий уровень углеводов.
Углеводы перевариваются рыбами значительно хуже, чем теплокровными животными. Степень их переваримости у рыб зависит от их строения. Простые сахара моносахариды усваиваются у форели гольца, карпа до 99 %. Для дисахаридов (сахароза, фруктоза) этот показатель варьирует у разных видов в пределах 52-73 %. Лактоза усваивается форелью на 60 %, осетрами – на 36 %. Полисахариды расщепляются и всасываются слабее сахаров. Лососевые рыбы переваривают 40 % углеводов, поступающих с кормом, карповые – 17-84 %. Сырая клетчатка лососевыми рыбами вообще не переваривается. Карп способен переваривать ее на 25-50 %. Расщепление сырой клетчатки происходит под действием ферментов, выделяемых панкреатической железой и кишечной микрофлорой карпа.
Потребность в липидах
Насыщенные и мононенасыщенные жирные кислоты могут синтезироваться в организме рыб из углеводородных остатков углеводов и белков. Полиненасыщенные жирные кислоты (ПНЖК) в организме рыб не синтезируются и должны поступать с пищей. У рыб, по сравнению с теплокровными животными, доля ненасыщенных жирных кислот в составе липидов значительно выше, что связано с относительно низкой температурой обитания. Рыбы, обитающие в разных условиях среды при уровнях различных температуры и солености, отличаются разным соотношением полиненасыщенных кислот. Содержание полиненасыщенных жирных кислот у морских рыб выше, чем у рыб, обитающих пресных водоемах. У пресноводных холодолюбивых видов рыб содержание полиненасыщенных жирных кислот выше, чем у теплолюбивых рыб.
Недостаток жиров и незаменимых жирных кислот приводит к нарушению физиологических функций организма, замедлению роста, ослаблению пигментации, некрозу лучей плавников, перерождению печени и почек, обводнению тканей и повышению смертности, снижению репродуктивной функции.
Потребности рыб в жире и белке взаимосвязаны. Жиры как источник энергии обладают протеин-сберегающим действием. При составлении рационов следует соблюдать соотношение уровня протеина с уровнем жира. Если это соотношение нарушено и в комбикормах доля протеина превышает оптимальное соотношение протеин/жир, то протеин будет расходоваться на энергетические нужды. При составлении рецептов комбикормов стремятся сократить использование белка для энергетических трат, сохраняя его для роста и обновления тканей. Для лососевых рыб при уровне протеина 50 % уровень жира должен достигать 15 %, а при уровне 30 % количество жира не должно превышать 5 %.
Потребность в минералах
Дается представление об особенностях усвоения минеральных веществ у гидробионтов, значимости микроэлементов и микроэлементов в минеральном обмене и функциях наиболее значимых элементов в питании рыб. Приводятся величины потребности отдельных элементов для некоторых объектов аквакультуры.
Минеральные вещества не являются питательными веществами, однако необходимы гидробионтам для нормального функционирования метаболизма. Особенность минерального питания рыб является поступление макроэлементов и микроэлементов организм не только с пищей, но и из воды осмотическим путем из воды через жабры, слизистые покровы ротовой полости и кожу. По количественному содержанию (более и менее 100 мг/кг) в живой материи их подразделяют на макро- и микроэлементы. В среднем общая сумма минеральных элементов в теле рыб составляет 2,5-8,0 %, из них доля макроэлементов составляет более 95,5 %. Потребность рыб в минеральных веществах составляет 4-5% от массы корма.
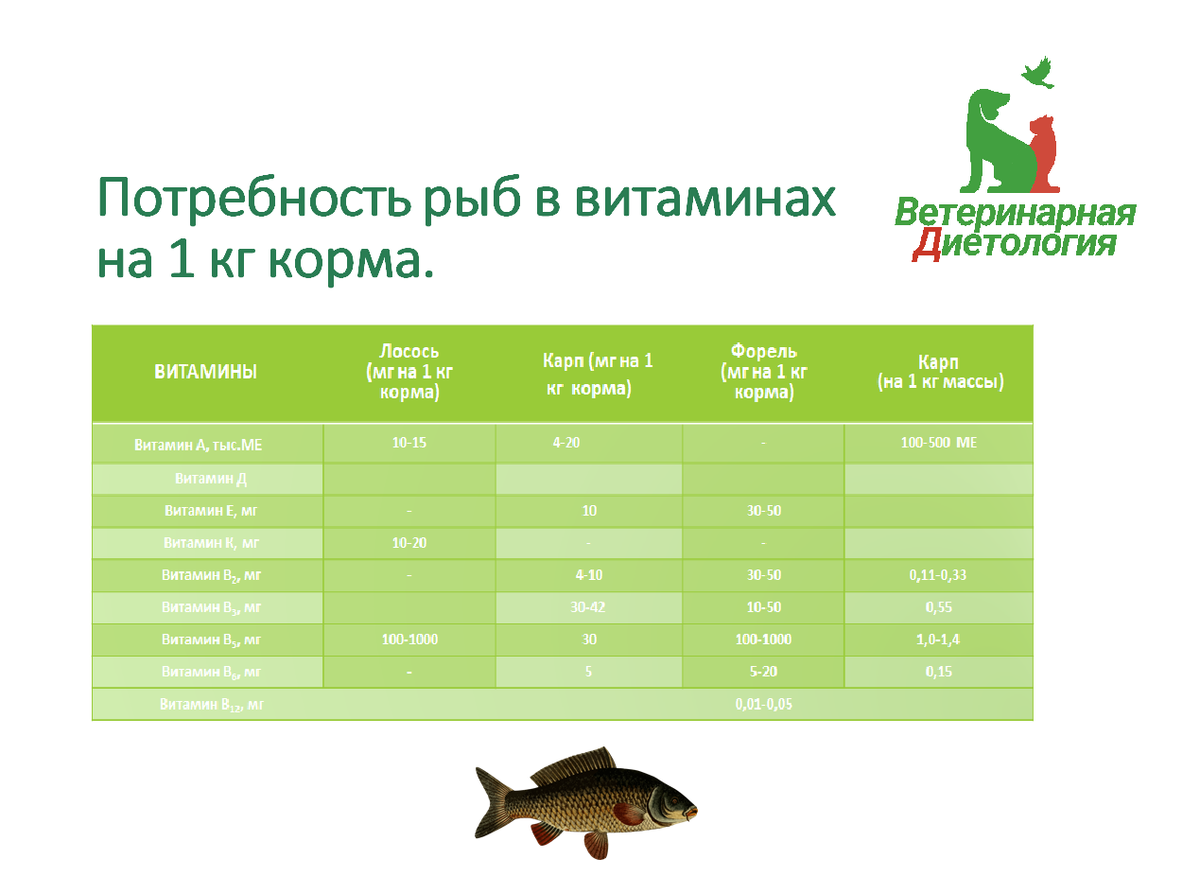
Потребность в витаминах.
Дается представление о значении витаминов в метаболизме рыб, описываются их функции в биохимических процессах в организме. Дается представление об антиоксидантах и их роли в искусственных кормах для гидробионтов.
Витамины – важнейшая группа незаменимых факторов. По химической природе они представляют собой низкомолекулярные, биологически активные органические вещества, необходимые для нормальной жизнедеятельности организма.
По физическим свойствам витамины подразделяются на жирорастворимые и водорастворимые.
Жирорастворимые витамины должны поступать с пищей. К жирорастворимым витаминам относятся А, Д, Е.
Витамин А (ретинол) принимает участие в обмене белков и минеральных веществ, оказывает влияние скелетообразование, плодовитость, зрение, обеспечивает нормальное состояние эпителия, является природным антиоксидантом. Для рыб провитамином А служат ксантофиллы (астаксантин, кантаксантин, зеаксантин), которыми богаты водные беспозвоночные. Наибольшее количество астаксантина содержат ракообразные являющиеся пищей рыб в молодом возрасте, а для многих и в течение всей жизни.
Всасывание витамина А и каротиноидов происходит в тонком кишечнике с участием желчи, обеспечивающей их эмульгирование. Основным депо в организме является печень. Отсутствие ретинола в пище тормозит рост рыб.
Витамин Д – группа веществ, объединенных под названием кальциферолы. Витамин Д способствует всасыванию кальция и фосфора в кишечнике, поддерживает их уровень в крови, активизирует переход из крови в костную ткань, непосредственно влияя на минерализацию скелета, улучшает усвоение магния. Дефицит его вызывает патологические изменения в мышечной и костной тканях. При дефиците его потребность в кальции и фосфоре увеличивается.
Витамин Е (ά-токоферол) представляет группу токоферолов. Витамин Е стабилизирует клеточные мембраны, предохраняя от окисления содержащие селен белки, подавляет самоокисление ненасыщенных жирных кислот, витаминов, регулирует развитие и функционирование половых желез, стимулирует процесс образования антител. Дефицит его вызывает патологические изменения в мышечной и костной тканях. При дефиците его вызывает нарушение целостности клеточных мембран, функций кроветворения и размножения, ожирение и некроз печени.
Витамин F представляет собой группу незаменимых полиненасыщенных жирных кислот линолевого, линоленового, арахидонового ряда. Принимает участие в синтезе жиров, обмене холестерина, способствует снижению уровня насыщенных жиров теле и предотвращает отложение холестерина в сосудах и артериях; обеспечивает нормальное состояние кожных покровов; стимулирует иммунную защиту организма; влияет на процесс сперматогенеза; участвует в образование простагландинов; во взаимосвязи с витамином Д содействует усвоению фосфора и кальция, которые необходимы для костной ткани.
Водорастворимые витамины (витамины группы В, С, Н).
Витамины группы В синтезируются главным образом, растениями, бактериями, дрожжами и входят в состав основных ферментов, катализирующих различные реакции белкового, липидного и углеводного обменов.
Витамин В1 (тиамин) является кофактором трех важнейших ферментов, участвующих в ключевых реакциях обмена углеводов – окислительном декарбоксилировании, в цикле трикарбоновых кислот и пентозофосфатном пути окисления углеводов. При его недостатке у рыб нарушается координация, снижается потребление корма.
Витамин В2 (рибофлавин) осуществляет реакции дегидрирования, входит в состав флавиновых ферментов, участвует в углеводном обмене, способствует образованию гликогена в печени.
Витамин В3 (пантотеновая кислота) является кофактором ферментов клеточного обмена.
Витамин В4 (холин) участвует в жировом обмене, входит в состав лецитина, поставляет метильные группы в реакциях метилирования.
Витамин В5 (никотиновая кислота) активизирует действие инсулина, влияет на уровень холестерина в крови и нормализует водно-солевой обмен.
Витамин В6 (пиридоксин) участвует в белковом и жировом обменах при переаминировании, декарбоксилировании и метилировании аминокислот улучшая использование незаменимых жирных кислот.
Витамин Н (биотин) является коферментом реакции карбоксилирования в липидном, белковом и углеводном обмене.
Витамин В8 (инозитол) входит в состав фосфолипидов, регулирует липидный обмен, предотвращая жировую дегенерацию печени.
Витамин Вс (фолевая кислота) В12 (цианкобаламин) стимулируют синтез гемоглобина, катализируют белковый и липидный обмен.
| Вложение |
|---|