Резюме
Для получения химер зародышевой линии птиц были использованы клетки кур, выделенные из трех источников: бластодермы, эмбриональной крови и гонад. Бластодермальные клетки выделяли из свежеснесенных яиц, клетки крови и гонад - из зародышей 13-14 и 28 стадий развития соответственно и культивировали в течение 15 суток. Впервые в качестве агента подавляющего развитие первичных зародышевых клеток был использован бусульфан для получения бластодермальных химер птиц. Только в случае использования бластодермальных клеток были получены половые химеры. Разработанный метод позволяет получение химер зародышевой линии птиц на уровне 70%, а выраженность данного признака варьирует в пределах от 4 до 61%.
Введение
Криоконсервация зародышевых клеток у птиц – способ сохранения генетического материала для улучшения сельскохозяйственной птицы в будущем и сохранения исчезающих видов. Для некоторых видов сельскохозяйственных птиц уже разработаны методы криоконсервации спермы, но консервация оплодотворенных яиц или эмбрионов, как это имеет место у млекопитающих, до сих пор не достигнута. Главным образом, это связано с крупными размерами яиц птиц и большим количеством желтка в них.
Восполнение данного пробела – основная цель многочисленных работ последних двух десятилетий, изучающих клетки ранних эмбрионов птиц. Предметом исследований в этих работах являются три типа клеток:
1. Бластодермальные клетки,
2. Эмбриональные стволовые клетки (ЭСК),
3. Первичные зародышевые клетки (ПЗК).
Не так давно была доказана тотипотентность первичных зародышевых клеток, которые являются предшественниками клеток спермы и яйцеклеток. Стволовые клетки, полученные из ПЗК, называются эмбриональными зародышевыми клетками (ЭЗК).
У птиц, в частности, у кур, на X стадии развития, что соответствует свежеснесенному яйцу, в бластодерме имеется немногочисленная (до 30-40 шт.) популяция клеток, обладающая тотипотентными свойствами. Эти клетки локализованы в центральной области бластодермы. В литературе отсутствуют данные о непосредственном выделении ПЗК на более ранних стадиях развития и их изучении.
Основная проблема, возникающая на пути интенсивного использования ПЗК в программах по сохранению генофонда и получению трансгенной птицы - это малое их количество при выделении. Для решения данной проблемы применяются различные приемы:
1. Концентрирование ПЗК в градиенте различных веществ.
2. Различные методы для уменьшения количества ПЗК эмбриона – реципиента: облучение ультрафиолетом [1], гамма-радиацией [2], лазером [3], химическая обработка бусульфаном [4], механическое вырезание зародышевого полумесяца [5].
3. Увеличение количества путем культивирования ЭСК и ПЗК. Очень важно обеспечить культивирование данных типов клеток в чистоте, т.е. не содержащими включений других типов клеток. Данное условие необходимо для точного контроля за результатами исследований по замораживанию и уровнем трансмиссии чужеродными генами при трансгенезе птиц. Кроме того, при этом отпадает необходимость проведения процедур по концентрированию нужной популяции клеток. Поэтому основное внимание уделяется именно этой проблеме, проблеме культивирования эмбриональных стволових клеток, обладающих тотипотентными свойствами.
Наиболее привлекательным и доступным средством подавления размножения ПЗК реципиентных эмбрионов является бусульфан, Bu (1,4-butandiol dimethanesulfonate). Бусульфан – алкилирующий агент, механизм действия которого основан на сшивании нитей ДНК, в результате которого нарушается процесс репликации молекулы наследственности. Считается, что в отличие от других алкилирующих веществ, Bu может оказывать действие на клетки, находящиеся в состоянии покоя. Время полужизни Bu составляет 10-12 часов. Он находит широкое применение в медицинской практике при лечении различных видов лейкемии. В многочисленных работах на млекопитающих и птице доказан супрессивный эффект бусульфана на развитие половых клеток [6,7,8].
Согласно литературным данным бусульфан используется в качестве агента для подавления размножения хозяйских первичных зародышевых клеток на стадии их миграции (48, 60 и 72 часа инкубации) к месту локализации гонад [9,10]. При применении сравнительно высоких концентраций от 25 до 125 мкг/яйцо достигается почти полная стерильность гонад обработанных зародышей. Имеются также сообщения о неэффективности применения концентраций меньше 400 мкг/яйцо [11], и довольно высоком уровне стерильности гонад при применении концентраций в пределах до 250 нг/яйцо [12]. Предлагались также различные способы растворения и доставки бусульфана к развивающемуся зародышу: растворение в диметилформамиде или диметилсульфоксиде и доставка при помощи кунжутного масла. Хотя авторы и отмечают довольно высокий индекс стерильности обработанных гонад (элиминация 2/3 половых клеток), однако, уровень трансмиссии зародышевой линии при инъекции донорских клеток увеличивалась всего лишь от 4 (в контроле) до 14% [13].
Следует отметить, что во всех случаях, когда применялся бусульфан, последующая инъекция донорских клеток производилась в кровяное русло зародышей 3-3,5 суточного возраста. Техника микроинъекций в кровяное русло на ранних стадиях развития значительно сложнее осуществить из-за частых случаев кровоизлияний и гибели зародышей, чем введение клеток в подзародышевую полость.
Таким образом, несмотря на положительный эффект, наблюдаемый во многих работах, следует отметить, что часто результаты действия Bu оказываются непредсказуемыми. Поэтому такие параметры, как продолжительность и место воздействия, концентрация Bu, стадия развития эмбриона, нуждаются в уточнении для получения стабильных результатов.
Материалы и методы.
Выделение бластодермальных клеток. Бластодиски выделяли из свежеснесенных оплодотворенных яиц при помощи кольца из фильтровальной бумаги [14]. Полученные эмбрионы дважды отмывали от желтка в растворе фосфатно-солевого буфера (ФСБ) (170 мМ NaCl; 3,4 мМ КС1; 4 мМ Na2HPO4; 1,8мМ КН2РО4; рН 7,2). Затем по 10-12 эмбрионов переносили в 1 мл ФСБ, содержащей 0,25% трипсина и 0,04% ЭДТА, и инкубировали 10 мин при 37°С, после чего пипетировали пастеровской пипеткой и центрифугировали 10 секунд при 1500 об/мин. Далее, осажденные клетки ресуспендировали в 1 мл питательной среды 199, содержащей 10% сыворотки плодов коровы. Суспензию клеток концентрировали центрифугированием 10 секунд при 1500 об/мин с последующим удалением 0,7 мл супернатанта, а затем клетки вновь ресуспендировали в оставшейся среде.
Выделение ПЗК для культивирования. Для получения первичных зародышевых клеток из крови использовали эмбрионы 13-14 стадии, для чего яйца инкубировали на протяжении 48-52 часов. От каждого эмбриона было получено по 2 - 4 мкл крови. Для выделения клеток крови в чашку Петри с питательной средой переносили кровь от 7-10 эмбрионов. Суспензию клеток отмывали путем центрифугирования (1000 об/мин) в течение 2 мин, а затем переносили в 2-3 лунки 24 луночного планшета для культивирования.
Для выделения ПЗК из гонад использовали эмбрионы 28 стадии развития. Для этого яйца инкубировали на протяжении 6 суток в стандартных условиях. Гонады 6-суточных эмбрионов выделяли и переносили в фосфатно-солевой буфер (рН 7,2) без ионов кальция и магния. В одном эксперименте выделяли гонады от 10-12 эмбрионов, которые помещали в центрифужную пробирку с теплым раствором трипсина (0,25%) и ЭДТА (0,05%). После инкубации с раствором в течение 5 мин. пипетировали пастеровской пипеткой до получения гомогенной суспензии. Действие трипсина инактивировали добавлением питательной среды с сывороткой. После отмывания полученную суспензию использовали для культивирования.
Культивирование клеток. Культивирование клеток гонад, выделенных из 6 суточных эмбрионов, проводили в питательной среде следующего состава: ДМЕМ+F12(HAM) (Sigma-Aldrich) (1:1) + 12 % сыворотки плода коровы (Пан-Эко, Москва) +56 мкМ меркаптоэтанола (Sigma-Aldrich). Клетки крови, выделенные из эмбрионов 13-14 стадии развития по Гамбургеру и Гамильтону [15], культивировали в среде, кондиционированной первичными клетками гонад. Для приготовления кондиционированной среды клетки гонад культивировали до достижения монослоя (3-4 суток культивирования), после чего среду частично заменяли свежим питательным раствором. Полученный супернатант очищали от клеточных включений путем центрифугирования при 3000 об/мин. 15 мин. Кондиционированную среду перед использованием разбавляли свежим питательным раствором в соотношении 1:3.
Клетки культивировали в лунках 24-луночного планшета в течение 10-15 суток.
Получение химер птиц. В качестве реципиентов использовали эмбрионы кур породы белый леггорн, которые являются гомозиготами по доминантному признаку белой окраски (II), а в качестве доноров - эмбрионы кур породы род-айленд красный гомозиготный по аллелю дикого типа в этом локусе (ii).
Донорские клетки инъецировали в подзародышевую область яиц-реципиентов микропипеткой (внешний диаметр 50-70 мкм) через круглое отверстие диаметром 0,7 см в скорлупе. В каждый эмбрион вводили 3-4 мкл суспензии, содержавшей 600-1000 донорских клеток. Отверстие в яйце закрывали кусочком тонкой пленки, которую приклеивали к скорлупе при помощи белка и затем сверху заклеивали кусочком лейкопластыря большего размера.
В качестве химического агента, который подавляет деление ПЗК эмбрионов-реципиентов, использовали бусульфан.
Результаты.
Для учета времени действия Bu при инъекции донорских клеток в эмбрион-реципиент был проведен эксперимент по влиянию кратковременного снижения температуры во время инкубации на выводимость яиц. Результаты эксперимента представлены в таблице 1.
Таблица 1. Влияние кратковременного снижения температуры во время инкубации на выводимость яиц.
|
Варианты |
Количество (шт.) |
Выводимость яиц (количество/%) |
|
Контроль |
27 |
23 (85,2) |
|
Вариант 1 |
20 |
13 (65,0) |
|
Вариант 2 |
16 |
5 (31,3) |
|
Вариант 3 |
18 |
14 (77,5) |
Контроль - свежие неинкубированные яйца.
Вариант 1- яйца на протяжении 5 суток прогревались до 37,8 0С по 2 часа, после чего были инкубированы при стандартных условиях до вывода.
Вариант 2- яйца на протяжении 5 суток прогревались до 37,8 0С по 2 часа, просверливали отверстие в скорлупе, заклеивали, после чего были инкубированы при стандартных условиях до вывода.
Вариант 3- яйца на протяжении 48 часов прогревались до 37,8 0С, после чего инкубировались на протяжении 35 часов при 33 0С, после чего были переведены в стандартные условия инкубации до вывода.
Вскрытие яиц в варианте 3 на 4-ые сутки инкубации показало, что эмбрионы развиваются с отставанием: в опыте - 17 стадия развития (53-64 часа), в контроле -21 стадия (84 часа).
На основе полученных данных в дальнейших исследованиях для обработки яиц бусульфаном была принята схема, согласно которой яйца - реципиенты прогревались в инкубаторе при 37,8 0С на протяжении 10-12 часов, что соответствует промежуточной стадии формирования первичной полоски, после чего в каждое яйцо вводилось рассчитанное количество Bu. Bu вводили микропипеткой в подзародышевую полость эмбриона в объеме 3-5 мкл жидкости. Яйца-реципиенты инкубировались с Bu не менее 24 часов (время активности бусульфана при 330С).
Бусульфан растворяли в диметилформамиде. Диметилформамид, будучи сильным растворителем, обладает пагубным действием на живые ткани, поэтому конечную его концентрацию доводили до 10% разбавлением питательной средой ДМЕМ.
Результаты опыта по влиянию разных концентраций Bu на эмбриональное развитие представлены в таблице 2.
Таблица 2. Влияние разных концентраций Bu на эмбриональное развитие.
|
Кол-во Bu на яйцо (нг/яйцо) |
Кол-во яиц (шт.) |
Перевод на вивод (кол-во/%) |
Выводимость яиц (кол-во/%) |
|
100 |
11 |
8 (72,7) |
3 (27,2) |
|
200 |
15 |
13 (86,6) |
7 (46,6) |
|
300 |
15 |
11 (73,3) |
8 (53,3) |
|
480 |
15 |
7 (46,6) |
6 (40,0) |
|
0 (контроль) |
18 |
18 (100) |
18 (100) |
Следует отметить, что кроме представленных в таблице концентраций были апробированы и другие варианты: 50, 70 и 100 мкг Bu на яйцо. Бусульфан растворяли в диметилформамиде или диметилсульфоксиде, который потом разбавляли подсолнечным маслом в соотношении 1:1. Ни один из этих вариантов не дал приемлемых положительных результатов.
Как видно из представленных результатов, выводимость яиц в группах, где инъецировались 100 и 200 нг Bu ниже, чем при более высоких концентрациях. Возможно, это связано с тем, что сама процедура вскрытия яиц оказывает значительное повреждающее влияние на эмбриональное развитие, что было замечено нами ранее [16].
В группах, где инъецировались 200, 300 и 400 нг бусульфана на яйцо у полученных цыплят в суточном возрасте определяли массу гонад. Результаты представлены в таблице 3.
Таблица 3. Влияние бусульфана на массу гонад.
|
Вариант |
Количество (шт.) |
Mасса (мг.) |
|
Контроль (самки) |
8 |
10,25±0,52 |
|
Опыт (самки) |
8 |
8,56±0,77 |
|
Контроль (самцы) |
9 |
9±0,91 |
|
Опыт (самцы) |
11 |
6,36±0,32 * |
* достоверная при P
В дальнейших исследованиях для получения химер была использована предварительная обработка зародышей Bu в концентрации 300 нг/яйцо. Результаты представлены в таблице 4. Фенотипический химеризм проявлялся лишь после инъекции бластодермальних клеток - 5 шт. (45,5%). У двух взрослых кур из пяти донорское оперение практически полностью вытеснило родительское во время постнатального периода. Кроме цвета оперения химеризм проявлялся также по окраске скорлупы яиц и форме гребня (рисунок 1).
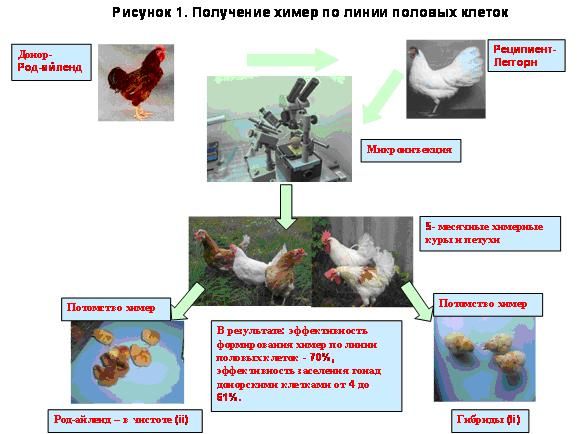
Таблица 4. Получение химер кур после инъекции донорских клеток.
|
Источник ПЗК |
Кол-во инкубированных яиц (шт.) |
Выводимость яиц (количество/%) |
Фенотипический химеризм (шт./%) |
% химеризма по окраске оперения |
|
Бластодерма |
38 |
11 (28,9) |
5 (45,5) |
5-100 |
|
Гонади |
81 |
13 (16,0) |
0 |
-- |
|
Кровь |
27 |
9 (33,3) |
0 |
-- |
Для анализа химеризма по линии зародышевых клеток химерные цыплята были выращены до половозрелого возраста. Химерные петухи были использованы для получения спермы и осеменения кур породы род-айленд красный, а молодки были осеменены спермой, полученной от петухов породы род-айленд красный. По результатам анализирующих скрещиваний был установлен процент химеризма по линии зародышевых клеток (процент химерных по линии половых клеток особей от общего количества выращенных до половозрелого возраста цыплят) и эффективность использования различных источников первичных зародышевых клеток: бластодермальные клетки, культивированные ПЗК из крови, и из гонад (соотношение донорского и реципиентного типов среди потомков химерных особей). Результаты анализа представлены в таблице 5.
Таблица 5. Результаты анализирующего скрещивания.
|
Источник донорских клеток: бластодермальные клетки |
||||||||
|
Пол птицы |
♀ |
♂ |
||||||
|
№ птицы |
1 |
2 |
3 |
4 |
5 |
1 |
2 |
|
|
Количество потомков |
46 |
24 |
24 |
32 |
20 |
41 |
3 |
|
|
Количество генотипов ii |
28 |
1 |
1 |
14 |
0 |
10 |
0 |
|
|
Количество генотипов Ii |
18 |
23 |
23 |
18 |
20 |
31 |
3 |
|
|
Доля половых химер, % |
60,87 |
4,16 |
4,16 |
43,75 |
0,00 |
24,39 |
0 |
|
|
Источник донорских клеток: культивированные клетки гонад |
||||||||
|
Количество проанализированных птиц |
5♀/2♂ |
|||||||
|
Количество потомков |
31 |
|||||||
|
Количество генотипов ii |
0 |
|||||||
|
Количество генотипов Ii |
31 |
|||||||
|
Доля половых химер, % |
0 |
|||||||
|
Источник донорских клеток: культивированные клетки крови |
||||||||
|
Количество проанализированных птиц |
5♀/2♂ |
|||||||
|
Количество потомков |
30 |
|||||||
|
Количество генотипов ii |
0 |
|||||||
|
Количество генотипов Ii |
30 |
|||||||
|
Доля половых химер, % |
0 |
|||||||
Как свидетельствуют представленные в таблице данные, при инъецировании бластодермальных клеток процент химеризма по линии зародышевых клеток составил 71,42% (5 из 7), а выраженность данного признака у различных особей варьировал от 4 до 61%. В то же время, при использовании для инъекции культивированных клеток гонад или крови не наблюдалось их включения в линию зародышевого пути. Это свидетельствует о том, что при использованных условиях культивирования данные типы клеток теряют свойство интеграции в линию половых клеток.
Обсуждение
При планировании экспериментов исходили из необходимости проведения процедуры микроинъекции донорских клеток в подзародыщевую полость, а не в кровяное русло. Поэтому в первом эксперименте мы выяснили, что на ранних стадиях развития (48 часовые эмбрионы) возможно снижение температуры инкубации –до 330С в течение 35 часов- без значительного снижения выводимости инкубируемых яиц - 77,5%. Контроль за развитием эмбрионов показала, что зародыши при таких условиях отстают в своем развитии на 24 часа. Это время можно использовать для химического воздействия на зародыш с целью подавления развития собственных ПЗК. Так как многие исследователи, работавшие с Bu, отмечали его тератогенный эффект [17], мы исходили из необходимости применения его минимальной концентрации. Такая концентрация может иметь эффект только в случае доставки химического агента в непосредственной близости от зародыша. Бусульфан инъецировали проколом центральной области бластодермы в подзародышевую полость. Минимальное количество подзародышевой жидкости на промежуточной стадии формирования первичной полоски должно было обеспечивать оптимальные условия для проявления алкилирующего эффекта Bu. Анализ влияния различных концентраций Bu на выводимость инкубируемых яиц показал сравнительно высокие результаты выводимости яиц (более 50%) при использовании концентрации 300 нг/яйцо, которая была использована в дальнейших экспериментах.
В более ранних исследованиях нами было выяснено (данные в печати), что при культивировании клеток крови (55 часов инкубации) в питательной среде, кондиционированной с клетками гонад, выделенных из 6 суточных эмбрионов, определенная популяция клеток садится на подложку культивирования и пролиферирует. Эта популяция клеток окрашивается при Шифф реакции в малиново–красный цвет, окраска характерная для первичных зародышевых клеток птиц. Поэтому наряду с бастодермальными клетками были испытаны культивированные клетки крови и гонад на способность формирования химер зародышевой линии. Как свидетельствуют приведенные в таблице 5 данные, только при использовании бластодермальных клеток, выделенных из свежеснесенных яиц, были получены половые химеры. Из 11 бластодермальных химер до половозрелого возраста выжили 7 особей: 5 курочек и 2 петушка. Следует отметить, что у петуха №2 сперма была значительно худшего качества, чем у первого, которая в начале состояла только из плазмы. Из 37 яиц, полученных от курочек породы род-айленд красный, осемененных этим петухом (возраст петуха превышал 7 месяцев), оплодотворенными оказались всего лишь 3. Вылупившиеся цыплята были гибриды (Ii), что свидетельствовало о том, что, несмотря на явно выраженную стерильность гонад, донорские первичные зародышевые клетки не заселили гонады эмбриона-реципиента.
Таким образом, разработанный метод получения химер зародышевой линии включает следующие этапы:
1. Прединкубация яиц-реципиентов 8-10 часов при 37,8 0С (промежуточная стадия формирования первичной полоски);
2. Химическая обработка яиц-реципиентов алкилирующим агентом бусульфан. Бусульфан вводится в подзародышевую полость в концентрации 300 нг/яйцо.
3. Инкубация яиц не меньше 24 часов при 30-32 0С – время действия бусульфана, на протяжении которого он теряет свою активность.
4. Инъекция донорских клеток в подзародышевую полость и инкубация до вывода при стандартных условиях инкубации.
Метод позволяет получение химер зародышевой линии птиц на уровне 70%, а выраженность данного признака варьирует в пределах 4 до 61%. Разработанный метод можно использовать для разработки режимов замораживания генетических ресурсов птиц.
1. Reynaud, G. Reproductive capacity and offspring of chickens submitted to a transfer of primordial germ cells during embryonic life [Текст] / Reynaud, G.//Roux’s Archives of Developmental Biology. -1976. -№179. –P. 85-110.
2. Thoraval, P. Somatic and germ-line chicken chimaeras obtained from Brown and White Leghorns by transfer of early blastodermal cells [Текст] / Thoraval P., Lasserre F., Coudert F., Dambrine G. // Poultry Science. -1994. -№73. –P.1897-1905.
3. Mims, M.F. Laser irradiation of the chick embryo germinal crescent [Текст] / Mims, M.F., McKinnel, R.G.//Journal of Embryology and Experimental Morphology. -1971. -№26. –P.31-36.
4. Aige-Gil, V. Sterilisation of avian embryos with busulphan [Текст] / Aige-Gil V., Simkiss, K. //Research in Veterinary Science. -1991. -№50. –P.139-144.
5. McCarry, J.R. Functional differentiation of chick gonads following depletion of primordial germ cells [Текст]/McCarry, J.R., Abbott, U.K.//Journal of Embryology and Experimental Morphology. -1982. -№68. –P.161-174.
6. Vick, L. Germ-line chimaeras can produce both strains of fowl with high efficiency after partial sterilization [Текст]/ Vick, L., Luke, G., Simkiss, K.// Journal of Reproduction and Fertility. -1993. -№98(2). –P.637-41.
7. Brinster, R. L. The effect of cells transferred into mouse blastocyst on subsequent development [Текст]/ Brinster, R. L. // Journal of Experimental Medicine. -1974. -№140. –P.1049-1056.
8. Ogawa, T. Recipient preparation is critical for spermatogonial transplantation in the rat [Текст]/ Ogawa, T., Dobrinski, I., Brinster, R.L.// Tissue & Cell. -1999. -№31. –P.461-472.
9. Reynaud, G. Action du busulfan sur La lignee germinal de L’embryo de poulet [Текст]/ Reynaud, G. //Extrait Bull. Soc. Aool. France. -1977. -№102. –P.417-429.
10. Reynaud, G. The effect of busulfan on the germ cell line of the quail embryo [Текст]/ Reynaud, G.// Arch. Anat. Microsc. Morphol. Exp. -1981. -№70. –P.251-258.
11. Hallett, J. S. The effects of busulfan on gonadal differentiation and development in Japanese quail (Coturnix coturnix japonica) [Текст]/ Hallett, J. S., Wentworth, B. C.// Poultry Science. -1991. -№70(7). –P. 1619-1620.
12. Furuta, H. Proliferation of exogenously injected primordial germ cells (PGCs) into busulfan-treated chicken embryos [Текст]/ Furuta, H. and Fujihara, N.// Asian Journal of Andrology. -1999. -№ 1(4). –P. 187-190.
13. Vick, L. Germ-line chimaeras can produce both strains of fowl with high efficiency after partial sterilization [Текст]/ Vick, L., Luke, G., Simkiss, K.// Journal of Reproduction and Fertility. -1993b. -№98(2). –P. 637-41.
14. Lucas, A. M. Atlas of Avian Hematology [Текст]/ Lucas, A. M., Jamroz,C.// Washington. D. C: U.S.Department of Agriculture, 1961.
15. Hamburger, V. A series of normal stages in development of the chick [Текст]/ Hamburger, V. and Hamiliton, H. L.// Journal of Morphology. -1951. -№88. –P.49-92.
16. Тагiров, М.Т. Вдосконалення методу реабiлiтацii оперованих яєць [Текст]/ Тагiров, М.Т. [та ін.]// Украiнска конференцiя молодих учених та аспiрантiв з питань птахiвництва: Тез. доп. - Харькiв, 1992. - С.4.
17. Swartz, W. J. Response of early chick embryos to busulfan [Текст]/ Swartz, W. J.// Teratology. -1980. -№21. –P.1-8.
Тагиров М.Т., Терещенко А.В., Терещенко Л.В. (Институт птицеводства УААН)