
Компостный червь Eisenia fetida (Savigny, 1926) является наиболее широко используемым человеком в коммерческих целях. Этот вид червя легко адаптируется к широкому спектру изменений окружающей среды и различному виду корму. Стратегия выживания этого вида червей также хорошо изучена в лабораторных условиях. Однако большинство научных данных по репродуктивности этого вида червей, полученных в условиях лаборатории, не соответствуют таковым при коммерческом крупно-масштабном вермикомпостировании, что и побудило провести данные исследования.
Коммерческие вермикомпостирующие компании и индивидуальные садоводы обнаружили, что характеристики выживаемости и репродуктивности отличаются существенно от таковых характеристик, опубликованных в научных работах для этого вида червей.
В данной работе изучали влияние факторов, которые могут повлиять на репродуктивность и выживаемость компостного червя E. foetida. Процесс вермикомпостирования в вермиконтейнерах и в буртах зависит от плотности популяции компостных червей. Зрелые, но непродуктивные особи продуцируют наименьшее количество копролита в день на одну особь, по сравнению с молодыми растущими особями и репродуктивными червями. Идеальная популяция - это активно размножающаяся популяция червей, которая дает максимальное количество конечного продукта – вермикомпоста или биогумуса. Идеальная популяция дождевых червей может быть функционально более «женской», то есть в составе, которой больше “женских” особей, где больше суперспариваемых особей и наибольшее число ювенильных, которые наиболее активно питаются. Существует только один способ получения идеальной популяции дождевых червей - это поддержание популяции, в которой черви функционируют как “женские” особи.
Предварительно было изучено влияние следующих факторов на сексуальный статус дождевых червей:
1) влияние биомассы;
2) влияние стресса;
3) влияние возраста;
4) необратимость полов;
5) изменение пола – принудительное или случайное;
6) влияние температуры;
7) влияние доступности корма;
8) имеются ли какие-либо общения у червей на химическом уровне, которые бы определяли пол.
В каждом опыте использовалось по 40 пар дождевых червей для статистической достоверности. Все черви были выращены отдельно, чтобы до начала экспериментов не происходило спаривания между червями, то есть в опыты брались “девственные” особи червей. Все черви были выращены при 25°С, за исключением опытов по влиянию различных температур. Всех червей кормили одним и тем же объемом корма на основе телячьего навоза (5 г), за исключением тех опытов, где тестировали доступность корма для червей.
Все пары червей для спаривания определялись с помощью статистических таблиц. Черви взвешивались до начала периода спаривания, в конце периода спаривания и еженедельно после спаривания. Все коконы отбирались еженедельно и подсчитывались. Каждая пара червей или индивидуально спариваемые или спариваемые в группе были зарегистрированы. Поведение наравномерного спаривания было определено для каждой пары или группы спаривания из полученных данных после завершения каждого эксперимента.
Влияние биомассы червей
В этих опытах была обнаружена прямая сильная корреляция между биомассой червей и полом функционирования. Черви с более высокой биомассой при спаривании были сравнимы со своими партнерами, имеющими на 100 мг меньшую биомассу в начале периода спаривания, неодинаковость особей спаривающейся пары червей показана на рис. 1. В группе червей, имеющей большую биомассу, “сверхспаривающихся” червей было 76%. Эти данные коррелируют с таковыми для других организмов. При более высоких значениях биомассы черви функционировали как “женские” особи.
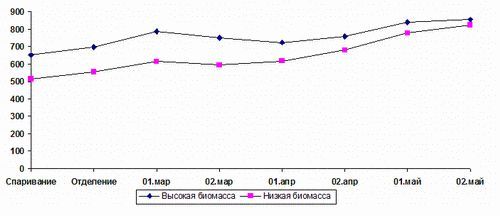
Рис. 1 Изменения биомассы в репродуктивный период у отдельных червей - сверспаривающихся и малоспаривающихся пар.
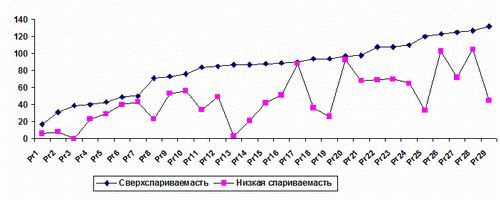
Рис. 2. Общее количество потомства, продуцируемое на одну особь для «сверхспариваемых» и «малоспариваемых» особей, определенных по данным для «сверхспариваемых» и «малоспариваемых» групп.
Влияние плотности популяции
Влияние плотности популяции на поведение животных и репродуктивную способность или производительность изучена для многих видов животных. Для компостного червя E. fetida было показано, что популяция червей с низкой плотностью усиливает свою репродуктивность - индивидуальные черви при отделении в контейнеры продуцируют большее количество коконов, чем черви, находящиеся в группе.
Спаривание червей изучали в группах по 2, 3 и 5 особей в этом эксперименте.
Группа из 2 червей. Пары особей служили контролями для определения критерия неравноспариваемости. В этой группе черви проявляли ожидаемый характер поведения неравноспариваемости.
Группа из 3 червей. Спаривались три червя, помещенные в один контейнер. У них обнаружен широкий диапазон в репродуктивности. Коконы, продуцируемые тройкой червей, отбирались и подсчитывались отдельно, а затем все сверхпродуценты были обозначены как «сверспариваемые», функционирующие как женские особи. Высокопродуктивные продуценты были обозначены как средние, а продуценты с низшей продуктивностью - как «малоспариваемые», функционирующие как «мужские» особи. Средняя группа была ближе к группе «сверспариваемых», показывая, что в группе из трех червей имелись два червя, функционирующих как женские особи и один червь как мужская особь. Эти данные требуют дальнейших исследований, так как популяционный стресс мог влиять на поведение троек червей в период спаривания.
Группа из 5 червей. Результаты этих опытов показали, что имелись очень большие различия в репродуктивности коконов у отдельных червей этой группы. В каждой «пятерке» червей имелись как особи с высокой репродуктивностью, так и низкой. Обнаружено, что три особи имели высокую репродуктивность (со свойствами «сверспариваемости») и два червя - низкую репродуктивность (со свойствами «малоспариваемости»).
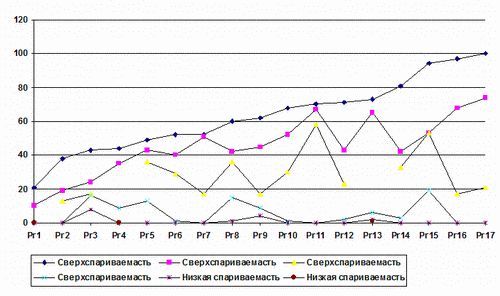
Рис. 3. Свойства неравномерности в популяции их пяти червей, спаривавшихся в течение 24 часов (количество потомства).
Популяционный стресс у дождевых червей в данном опыте мог быть обнаружен таким же способом, как и в группе из трех особей, но результаты опытов имели ту тенденцию - появлялось больше особей с “женскими” функциями, чем особей с “мужскими “ функциями. Если популяционный стресс является фактором, то, как было показано Meyer (1998) в буртах (горизонтальное пространство) не было преимущества по репродуктивности, а в контейнерах (вертикальное пространство) наблюдалось отрицательное влияние. Эти результаты показывают, что «сверспаривамые» черви являются индикатором того, что популяционный стресс не влияет на репродуктивность отдельных червей.
Влияние возраста
У некоторых организмов стратегия репродуктивности имеет следующую тенденцию: старые животные превращаются в женские особи с более высокой биомассой, а молодые становятся мужскими особями с меньшей биомассой. В этом эксперименте мы определяли влияние возраста червей на феномен неравнопариваемости. Были созданы две группы из «девственных» червей. Разница в возрасте составляла 4 месяца. Неравноспариваемость была четко доказана в этих экспериментах. Существует тесная корреляция между возрастом червей и сверхспариваемостью. Так, 83% червей старшей возрастной группы являлись сверхспариваемыми.
Необратимость полов
Факт, что специфические функции при сверхспаривании, могут быть объяснены переходом от сверхспариваемости к малоспариваемости и наоборот. Этот феномен не существует у других организмов. Были использованы черви от предыдущих опытов, у которых преобладало поведение неравноспариваемости. Малоспаривамые черви спаривались с малоспариваемыми, а сверхспариваемые со сверхспариваемыми особями.
Эти результаты снова показывают наличие у червей неравноспариваемости. Это означает, что один партнер изменяет сексуальное предпочтение. В тех экспериментах, где малоспариваемые спаривались с малоспариваемыми червями, один имел поведение малоспариваемости. В эксперименте, где сверхспариваемые черви спаривались с сверхспариваемыми, в один приобретал свойства малоспариваемости. Это показывает, что этот вид компостных червей и, вероятно, и другие виды дождевых червей могут изменять характер репродуктивности с целью выживания вида.
Изменение пола - принуждение или случайность?
В этом эксперименте изучали фактор, который мог бы влиять на изменение сексуальной предпочтительности. Черви из других экспериментов, где у них наблюдалась неравноспариваемость, помещались в контейнер для спаривания с молодыми (девственными) особями. Спаривание происходило по методу, предварительно описаному. Сверхспариваемые черви из других экспериментов спаривались с молодыми «девственными» червями, а малоспариваемые черви спаривались с «девственными» червями. В обоих опытах была обнаружена неравноспариваемость у червей. В опытах со слабоспариваемыми червями 52% червей превращались в червей со свойствами сверхспаривамых. Это могло произойти в результате того, что биомасса у слабоспариваемых червей была выше, так как они были более крупными и жирными. В опытах со сверхспариваемыми червями, которые имели биомассу близкую к таковой «девственных» червей, результаты также показали неравноспариваемость. Эти результаты не показывают на стабильность или изменение их сексуального поведения.
Влияние температуры
Известно, что температура влияет на репродуктивность многих видов дождевых червей. Для червей вида E. fetida, который обладает высокой толерантностью к температурным изменениям, показано, что температура ниже 15°С изменяет скорость роста, но увеличивает репродуктивность. В наших исследованиях червей, выращенных при температуре 15°С спаривали с червями, выращенными при 15°С, червей, выращенных при 25°С, спаривали с червями, выращенными при этой же температуре, а червей, выращенных при 15°С, спаривали с червями, выращенными при 25°С. Результаты опытов показали, что свойство неравноспариваемости при различии в биомассе двух групп червей не зависело от температуры. Эти результаты также говорят о том, что репродуктивность червей не изменялась при разнице температур в 10°С.
Влияние доступности корма
Пища играет важную роль для репродуктивности у большинства видов. Для некоторых видов доступность корма может изменять сексуальное поведение. Это было доказано, например, для крыс, хламидомонад, мух и других видов. В наших опытах по изучению влияния пищевого стресса производили замену пищевых субстратов, которые смешивали с вермикулитом, причем объем пищи был одинаковым, но пищевая ценность - разная. Известно, что дождевые черви могут рециклировать пищевой субстрат, в котором они обитают, даже при крайне неблагоприятных условиях. Исследовали сексуальное функционирование червей в зависимости от доступности корма в период, предшествующий спариванию, и во время периода спаривания.
Группа 1. Червей выращивали в смеси, содержащей 60% пищевого субстрата + 40% вермикулита. Червей спаривали в 100%-ном пищевом субстрате. Результаты показали неравноспариваемость. Но не было обнаружено доказательств того, что пищевой стресс у червей в период до их спаривания не вызывал свойств сверхспариваемости или малоспариваемости спаривания при их спаривании.
Группа 2. Черви, выращенные в среде из 60% пищевого субстрат + 40% вермикулита, затем спаривались в 60% -ном пищевом субстрате. При этом у червей наблюдались различия в половом поведении. Но не имелось доказательств того, что пищевой стресс у червей в период до спаривания мог вызвать сверхспариваемость или малоспариваемость в период спаривания.
Группа 3. Черви, выращенные в 100%-ном пищевом субстрате, спаривались с червями, выращенными в 100%-пищевом субстрате (контроль). В данном случае наблюдался эффект неравноспариваемости. Результаты данных опытов показывают, что пищевой стресс исключен у червей до спаривания и не изменяет сексуальное функционирование при спаривании и после спаривания.
Влияние хемокоммуникации
Большинство случаев хемокоммуникации у беспозвоночных происходит в результате выделения химических веществ (феромонов) самцами и самками. В наших опытах изучали влияние хемокоммуникации на сексуальную привлекательность или поведение у «девственных» червей. Если один из червей имел сексуальное поведение со свойствами сверхспариваемости (выживание вида) во время периода спаривания, то это должно быть сообщено другим червям, находящимися на его территории. В этом эксперименте один червь был перемещен без субстрата, в котором он обитал, в другой контейнер к другому червю. Не было обнаружено влияния на сексуальное поведение другого, хотя неравноспариваемость имела место.
Заключение
Факторы, которые могли влиять на сексуальную привлекательность E. fetida являются следующие: биомасса, плотность популяции и возраст. Между биомассой, качеством и количеством пищи имеется прямая связь, поэтому коммерческое вермикомпостирование должно уделять внимание уровню питания дождевых червей.
Плотность популяции должна контролироваться при коммерческом вермикомпостировании для того, чтобы достичь оптимальных результатов. Эти два фактора должны контролироваться и регулироваться для того, чтобы иметь в популяции дождевых червей оптимальное возрастное соотношение для получения наилучших результатов.
Автор(ы): W.J.Meyer, WormCo, PO Box 1361, Postchefstroom 2520, North West, South Africa
СП "Ecorosteh" Ltd.